Lipid Peroksidasyon
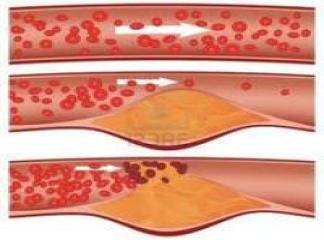
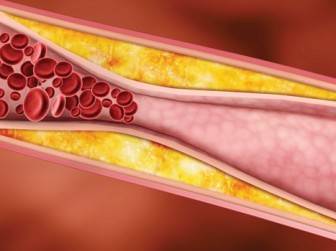
Lipid Peroksidasyon : Genelde hücre plasma membranı ve lipoprotein fosfolipid doymamış yağ asitlerinin peroksidasyonu, oksijen (O2".) ve hidroksil (OH.) radikalleri ile başlar. Lipid peroksidasyon zinciri transizyon metallerinin (örneğin Fe2+) katalitik etkisi ile birbirini takip eder. Pek çok ara ürünler, nihai maddeler [peroksitler, aldehidler, alkil radikalleri, volatil hidrokarbonlar (pentan ve etan) ve MDA] den oluşurlar. Pentan n-6 doymamış yağ asitlerinin (C18:2, C20:4, C22:4) etan ise n-3 doymamış yağ asitlerinin (C18:3, C20:5, C22:5 ve C22:6) peroksidasyon son ürünleridir. Volatil nihai ürünler organizmayı ekspirasyon esnasında terkederler. Lipid peroksidasyon seyrinde oluşan ara radikaller (R=lipid radikal, RO=lipid alkoksi radikal, ROO=Lipid peroksi radikal) yeni peroksidasyon reaksiyon zincirlerini harekete geçirirler ve dolayısıyla bir otokataliz reaksiyon zinciri başlamış olur.
Bu lipid hidroperoksid (ROOH) olayının başlamasına neden olan hidrojen atomu başka bir doymuş yağ asidi molekülü ile tutulursa oto oksidasyon zinciri önlenebilir. Böylesi bir molekül (antioksidan), lipid peroksit (ROO.) deki serbest elektronu tu- tar.Bu olaya glutatyon redüktaz (GRed) sistemi iştirak eder ve dolayısıyla otokatalitik reaksiyonun kesilmesine önemli katkıda bulunur. Endotel hücresinin pek çok önemli fonksiyonları membran doymamış yağ asitlerinin peroksidasyon ürünlerinden zarar görür. Bunlara ek olarak bu peroksidasyon ürünleri fosfolipaz A ve C için de son derece uygun substratlardır. Endotel de dahil değişik hücre membranlarınm gördükleri zararları şöylece sıralıyabiliriz:
(1) membran yağ asitleri kompozisyonundaki değişiklikler ve özellikle doymamışyağ asitlerinin azalması,
(2) membran permeabilitesindeki değişiklikler düşük ve yüksek moleküllü makromolekülere karşı perméabilité artışı, iyon kanallarındaki değişmeler ve membran sızıntısı,
(3) membran akışkanlığındaki değişmeler,
(4) membran proteinlerinde inaktivasyon, enzimatik hidroliz ve protein denaturasyonu.
Peroksidasyo- nun ara ürünlerinden 4-Hidroperoksi-none- nal, 4-Hidroksi-2-alkenallar ve MDA değişik enzimlerin sülfidril grupları için kuvvetli re- aktivatordürler. Hücre membranmdaki bu değişiklikler immün sistemde ve reseptör dağılımında belirgin bozukluklara neden olur.Oksidanlar, Kalsiyum ve Endotel ha- rabiyeti: Kalp dokusu üzerinde yapılan çalışmalar gösterdi ki lipid peroksidasyonu ürünleri kalsiyum metabolizmasını belirgin bir biçimde bozmaktadır. Bu gibi durumlarda en belirgin nokta hücrede kalsiyum birikimidir. Böylesi bir durum kalsiyum bağımlı fos- folipazların, kalsiyum bağımlı protein kinaz- ların, kontraktil elementlerin, kalsiyum ATP'azların kuvvetle aktivasyonuna neden olur ve ayrıca mitokondrilerde kalsiyum birikimine yol açar. Buna benzer kalsiyumla ilgili hücre harabiyeti endotelde daha az görülür. Yapılan az sayıda deneyler göstermiştir ki endogen oksidanların aktivasyonu (ksantin oksidaz-ksantin keneti) domuz pulmoner arter endotelinde kalsiyum aracılığı ile membran permeabilitesini bozuyor. Bu etki kalsiyum kanal blokürleri ve kalsiyum şelatörleri ile antagonize oluyor. Kalsiyum ionoforu A 23187 ise bu etkiyi potansiye ediyor.Lipid peroksidasyonu üzerine diyet faktörlerinin etkisi: Diyetle ilgili pek çok faktörler lipid peroksidasyonu hızını etkiler.
1. Peroksidasyon substratı olarak membran fosfolipidlerinde lokalize olan doymamış yağ asitleri, trigliseridler veya esterifiye olmamış doymamış yağ asitlerinden daha uygundurlar. Başka deyimle membran fosfoli- pidlerine bağlı olanlar çok daha kolay perok- sidasyona uğrarlar. Bunun olası nedeni oksijenin biyomembranlarda daha fazla çözünürlük kazanmasmdandır.
2. Poliansature fosfolipid molekülünün n- 3 serisi, 5 veya 6 çift bağ ihtiva eden poliansature yağ asitleri, n-6 serisi (C18:2, C20:4) ne nazaran peroksidasyona daha çok maruz kalırlar. Gerek hücre ve gerekse organelleri-nin membranları diyetle kolayca etkilenirler. Nitekim balık yağından zengin diyetle beslenen sıçanların karaciğer hücrelerinin endop- lasmik membranına C22:6 n-6 poliansature yağ asidinin enkorpore olması bu membramn in vitro koşullarda lipid peroksidasyona olan duyarlılığını artırıyor.
3. Yüksek sıcaklıkta dağlanan yağlarda lipid peroksidasyon ürünleri fazlaca oluşmaktadır. Deneysel olarak gerek miyokard ve gerekse damar duvarında lipid peroksidasyon birikimi ve buna bağlı anormallikler gösterilmiştir. Tavada dağlanmış yağla beslenen sıçanlarda karaciğerde lipid peroksidasyon ürünlerinin toplandığı gösterilmiştir.
4. Katalist metallerin varlığı in vitro poliansature yağ asitlerinin peroksidasyonunu hızlandırmaktadır. Bunlar arasında Fe2+ son derece kuvvetli bir katalitik metaldir. Fare karaciğeri mikrozomal membranlarmdaki lipid peroksidasyon ölçümleri yapılmıştır. Daha önce 5-10 mg Fe-dekstran kompleksi zer- kedilerek hem molekülüne bağlı olan ve olmayan Fe düzeyi artırılmış hayvanlardan elde edilen fraksiyonlarda lipid peroksidasyonu kontrol hayvanlarınkine nazaran anlamlı olarak artmış bulunmuştur. Kalpte iskemi re- perfüzyon harabiyetlerinde ve fonksiyonel bozukluklarında Fe şelatörlerinin doku koruyucu etkisi gösterilmiştir.
5. Vitamin E (Tokoferol)’nin antioksidan etkisi çok iyi bilinir.
Bu vitamin, margarinlerin içine havaya maruz kaldıklarında peroksidasyona uğramalarını önlemek için yeterli miktarda eklenir. Klinikte oksidatif stres süreci sonucu oluşan patolojik durumları tedavi için kullanılır. Diyetteki vitamin E büyük ölçütte karaciğer, az miktarda da kalp ve böbrek hücre membranlarında toplanır. Vitamin E kan-beyin engelini geçemez. Bu bakımdan beyindeki etkisi bilinmemektedir. Bu vitamin sarkoplasmik retikulum ve mitokondria membranmda toplanır. Plasma membran düzeyi düşüktür. Diyette herhangi bir manipü- lasyonla membranlardaki vitamin E düzeyini artırmak mümkün değildir. Diyete günlük ihtiyacın 10.000 katını ilave etmek dokulardaki vitamin E konsantrasyonunu artırabilir. Fakat bu lipofilik vitaminin membrandaki solü- bilitesi son derece zayıftır. Eğer diyette poliansature yağ asitlerinin düzeyi yüksek ise günlük vitamin E ihtiyacı da buna paralelolarak artar. Fakat balık yağı dışında diğer bitkisel poliansature yağ asitlerini içeren kaynaklarda vitamin E düzeyi de yüksektir.
Bu nedenle bitkisel kaynaklardan alman poliansature yağ asitlerinin peroksidasyonu mevcut vitamin E ile önlenmiş olur. Malnüt- risyon, alkolizm ve kistik fibrozise bağlı ma- labsorpsiyon hallerinde vitamin E alınımı azalır ve dokularda lipid peroksidasyon ensi- dansı yükselir. Vitamin E nin endotel hücresiyle ilişkisi de son derece önemlidir. Polimorf nüveli lökositlerin endotele yapışmasını vitamin E kuvvetle inhibe eder. Ayrıca endotel hücresinin PGI2 yapmasını stimüle eder ve endotelin korunmasını sağlar. Karotenlerin de (beta-karoten ve benzeri poliprenler) lipofi- lik serbest radikal yakalayıcılarıdır. Bitkilerde özellikle klorofil ve ışık etkisi ile oluşan serbest radikaller için tuzaktırlar. Ancak bunların in vivo antioksidan etkileri hakkm- daki bilgilerimiz fazla değildir. Askorbik asit (Vitamin C) beta-tokoferol rejenerasyonunda önemli bir uyarandır. Pro ve antioksidatif etkisi bu şekilde izah edilmektedir. Ancak çok önemli bir nokta, suda eriyen bir madde olan askorbik asidin nasıl ve hangi mekanizma ile lipofilik biyomembranlarda etkili olduğu izaha açıktır. Muhtemelen bu vitminin prooksi- datif etkisi ferritin ile Fe arasındaki etkileşmeyi modüle ederek olmaktadır.
Fazla Fe yüklenmiş hastalarda askorbik asit yüksek miktarda alındığında önemli zararlı etki ortaya çıkabilir. Zira bu vitamin ferritin-Fe3+ kompleksinden Fe2+ oluşumunu artırır. Daha önce de bahsedildiği üzere Fe2+ son derece kuvvetli bir katalitik metaldir ve peroksidas- yonun artışına neden olur.6. Serbest radikalleri tutan enzimlerin ak- tivitelerindeki artış son derece önemli bir koruyucu mekanizmadır. Diyette bulunan eser elementlerden selenyum (Se) glutatyon pe- roksidaz (GPer) enziminin optimum aktivite- si için gereklidir. Diyette Se yetersizliği nedeniyle kanser gelişebilmektedir. Keza Se yetersizliği Çin'de Keşan bölgesinde çok yaygındır ve çocuklardaki ağır kardiyomiyopatilerin nedeni olarak bilinmektedir. Finlandiya'da bir bölgede görülen iskemik kalp hastalıklarının nedeninin bu bölgedeki diyetle alınan selenyum yetersizliği olduğu gösterilmiştir.
Diyete selenyum ilavesiyle literatüre "Keşan Hastalığı" olarak girmiş olan yaygın patolojikdurum ve iskemik kalp hastalıkları başarı ile önlenebilmiştir. Se yetersizliği ile endotelde lipid peroksidasyon metabolitlerinin arttığı, malonildialdehid biriktiği, PGI2 sentezinin azaldığı ve bu durumların diyete Se ilavesi ile önlendiği gösterilmiştir. Se ilaveten optimal glutatyon peroksidaz aktivitesi için indirgenmiş glutatyona (GSH) da ihtiyaç vardır. Glutatyon bir tripeptiddir. Yapısı Gama-Gluta- mil-Sistein-Glisin'dir. Bunlardan sistein se- mi-esansiyel bir amino asittir. Organizmada ya metiyoninden sentez edilir veya diyetle alınır. Eğer yüksek düzeyde ihtiyaç hasıl olursa diette yüksek miktarda metiyonin alınması gerekir. Fakat N-Asetil sistein bu amino asidin yetersizliğinde en önemli kaynaktır. Deneysel olarak iskemi/reperfüzyon hasarlarında, tiyol havuzunun azaldığı durumlarda ve açlıkta oluşan protein yetersizliğinde ve bazı hepatotoksik ilaçlarla zehirlenmelerde GSH düzeyi ve aktivitesi azalır. Bu durumlarda N-Asetil sistein önemli bir anti- dot olarak kullanılır.
İLGİNİZİ ÇEKEBİLİR